تطور الحوت والأدلة العلمية الثابتة علي حدوثه (1)
كان موضوع تطور الحيتان والثدييات المائية موضع للجدل في فترات تاريخية سابقة، وكانت أحد إدعاءات الخلقيين ومعارضي التطور الشهيرة هي نفي تطور الحوت بسبب عدم وجود أي حفريات إنتقالية توضح الخط الزمني لتطور الحوت من الثدييات الأرضية.
ونقتبس في هذه النقطة ما قاله أحد أشهر الخلقيين (دوان جيش) من معهد أبحاث الخلق ..
Duane Gish of the Institute for Creation Research (ICR)
"There simply are no transitional forms in the fossil record between the marine mammals and their supposed land mammal ancestors."(Gish 1985: 78-9).
"لا يوجد ببساطة أي صور إنتقالية للسجل الحفري بين الثدييات المائية وما يفترض من أسلافها الثدييات الأرضية."
* بالطبع كان لسنين طويلة هناك نقص واضح في الحفريات الإنتقالية التي يمكنها تأكيد حقيقة تطور الحوت من الثدييات الأرضية لكن ذلك تغير بشكل قاطع مع الإكتشافات الحديثة في السنين الأخيرة.وقبل الدخول في نفي الإدعاء السابق وبسط الأدلة العلمية الحديثة علي حقيقة تطور الحوت نبدأ بسؤال أساسي.
ما هو الحوت ؟
الحوت أولاً وقبل كل شيئ هو (ثدي) فقّاري من ذوات الدم الحار؛ والتي تستخدم الميتابوليزم (الأيض) العالي داخلها لإنتاج الحرارة والحفاظ علي درجة حرارتها الداخلية. الإناث منها تحمل الصغار الذين يرضعون من غددها الثديية. علي الرغم من أن الحيتان الذكور البالغة غير مغطاة بالشعر فهي تكون مغطاة بالشعر بشكل مؤقت أثناء فترة الجنين وبعضها يبقي عندها شعيرات إستشعار حول فمها. هذا الخواص لا شك أنها خاصة بالثدييات.
ومع كونه من الثدييات لكنه يتميز بمجموعة من الخواص الفريدة والتي لا يشارك فيها الثدييات الأرضية، بل وبعضها حتي لا يشاركها مع أمثاله من الثدييات المائية مثل sirenians(manatees and dugongs) وأيضاً
pinnipeds (seals, sea lions, and walruses).
علي سبيل المثال فأجسام الحيتان streamlined (وتعني في علم ديناميكا الموانع بشكل مبسط أن تشكل الجسم يكون في وضع بحيث يكون مقاومة الهواء لحركته أقل ما يمكن) وهذه الأجسام سميكة ودائرية علي عكس أجسام الأسماك المستطيلة والرفيعة. ذيل الحوت هو أفقي وغرضه الأساسي هو دفع الحوت أثناء الحركة في الماء.
زعنفة الظهر مشدودة بأنسجة ربط وسمينة وبدون أي عظام لتدعيمها.
الزعانف الجانبية للحوت التي تسمح له بالقلب مكونة من عظام ذراع مسطحة وقصيرة، عظام رسغ مسطحة تشبه القرص وعدة أصابع مستطيلة.
عظام الرقبة للحوت قصيرة وعلي الأقل مدموجة بشكل جزئي في كتلة عظمية واحدة. الفقرات بعد الرقبة متعددة وتشبه بعضها البعض بشكل كبير؛ الأجزاء العظمية التي تربط الفقرات ضئيلة جداً بشكل يدعم مرونة هائلة لظهر الهوت وقوة دفع كبيرة لذيله في الماء.
وصلة الكوع تعد تقريباً غير متحركة بما يجعل الزعنفة جامدة.
في حزام الكتف، لوح الكتف مسطح ولا يوجد ترقوة. بعض الأنواع من الحيتان مازالت لديها حوض vestigial (أي باقي فيها نتيجة للأصول التطورية للنوع مع فقدانه وظيفته الأصلية وهي دعم الحركة علي الأرض) والبعض لديه أطراف خلفية – أيضاً vestigial- داخلية مصغرة ولاوظيفة لها بالنسبة للحوت.
القفص الصدري متحرك؛ وفي بعض الأنواع منفصل بالكامل عن العمود الفقري مما يسمح للصدر للتمدد بشكل كبير في حالة تنفس الحوت وأيضاً يسمح للصدر بالإنضغاط العميق في حالة غوص الحوت في أعماق كبيرة تحت الماء.
والجمجمة أيضاً لديها من الخواص الفريدة ضمن الثدييات. الفك ممتد للأمام مما يعطي الحيتان رأسه الطويلة
والمميزة؛ والعظمتين الأماميتين للفك العلوي(the maxillary and premaxillary) تشكلت بشكل تلسكوبي للخلف بشكل يغطي أحياناً قمة الجمجمة. هذا التكوين العظمي هو سبب كون فتحة الأنف للحوت أن تكون في قمة الجمجمة؛ ومحركاً المخ والجهاز السمعي لمؤخرة الجمجمة. الodontocetes(الحيتان ذوات الأسنان) لديهم فتحة نفخ واحدة والmysticetes (البالين) لديهم فتحتان نفخ.
علي عكس باقي الثدييات، الحيتان ليس لديها غدد دمعية، غدد جلدية أو حاسة للشم. قدرتهم علي السمع حادة لكن بدون أي فتحات خارجية. السمع يحدث نتيجة إهتزازات تنتقل لعظمة ثقيلة تشبه الصدفة تشكلت نتيجة إندماج لعظام الجمجمة (the periotic and auditory bullae).
تلك هي أهم الخصائص المميزة للحوت. بعضها يوضح بشكل ظاهر خصائص حاصلة نتيجة تكيف النوع مع بيئته البحرية؛ بعضها يربط الحيتان بأسلافها الأرضية. والبعض يظهر آثار من أصلها والذي يربطها بأسلافها الأرضية. من كل ذلك يمكننا أن نشكل الطريق الذي سارت فيه عملية تطور الحوت من السلف الأرضي إلي الحوت الحديث الذي يعيش في أعماق المحيط.
الدليل علي تطور الحوت ؟
الآن إلي الجزء الأكثر تشويقاً وأهمية في هذه المقالة؛ وهو الدليل علي إنحدار الحوت من ثدييات أرضية و سنقسمه هنا إلي 10 أقسام ..
1- paleontological (ويختص بالسجلات من الحفريات).
2- morphological (الأدلة المأخوذة من المقارنة الشكلية التشريحية بين حفريات الحيتان والثدييات الأرضية الحالية).
3- molecular biological (الأدلة المأخوذة من علم البيولوجيا الجزئية ودراسة الحمض النووي).
4- vestigial (ويختص بالآثار التطورية داخل الحيتان من سلفها الأرضي وفقدت –أو تغيرت- وظيفتها الأصلية).
5- atavism (ويختص بالخصائص الغير أصيلة للحيتان ومرتبطة بأصوله الأرضية وتظهر في تشريحه حتي بعد الولادة).
6- embryological (ويختص بدراسة علم الأجنة وما يتعلق بدراسة تطور الحوت منه والدليل علي حدوثه).
7- geochemical (ويختص بمدي توافق الأدلة علي تطور الحوت من ثدييات أرضية من الناحية الجيولوجية مع المحيط التي تواجد فيه من ناحية كيمائية).
8- paleoenvironmental (ويختص بمدي توافق الأدلة علي تطور الحوت من ثدييات أرضية من ناحية السجلات الحفرية المتوفرة وتوافقها مع البيئة المحيطة لكل إنتقال للحفريات من أقدم لأحدث من ناحية بيئية).
9- paleobiogeographical (ويختص بمدي توافق الأدلة علي تطور الحوت من ثدييات أرضية من ناحية السجلات الحفرية المتوافرة مع التنوع الجغرافي وشكل إنتشار الحفريات فيه).
10- chronological (ويضع جميع الأدلة السابقة في إطار زمني متدرج من الأقدم للأحدث متوافق مع ما تطرحه نظرية التطور من حقيقة إنحدار الحيتان من الثدييات الأرضية).
1- الدليل الحفري (Paleontological evidence)
الدليل الحفري يأتي من دراسة تسلسل الحفريات من الثدييات الأرضية بشكل تدريجي لأشكال تشبه الحيتان حتي الظهور النهائي للحيتان الحديثة. علي الرغم من أن الحيتان المتقدمة Archaeocetes لديها من التنوع والتفرع ما هو أكبر من المساحة المتاحة للحديث عنها هنا، لكن الأمثلة التي نطرحهها هنا توضح بوضوح الإتجاه الذي نود شرحه. وأيضاً علي الرغم من أن هناك تحت رتبتين للحيتان الحديثة الآن (Odontocetes and Mysticetes)، ففي هذه المناقشة سنكتفي فقط بدارسة تطور الحوت ككائن ثديي مع عدم الدخول في تفاصيل تشعبه إلي أكثر من رتبة حديثاً.
Sinonyx

Sinonyxjiashanensis (المتحف الوطني للطبيعة والعلوم، طوكيو اليابان)
سنبدأ بالSinonyx، وهوmesonychid بحجم ذئب تقريباً ((الmesonychid هي ذوات حوافر ungulate بدائية من الرتبة Condylarthra (رباعية الأطراف) والتي بدورها كانت الأصل للأنواع بعدها من ذوات الحوافر مثل الartiodactyls(وهي أنواع زوجية الأصابع أي تمشي بوزنها علي أصبعين الثالث والرابع مثل الحصان) والperissodactyls (وهي أنواع فردية الأصابع أي تمشي بوزنها علي أصبع أو ثلاثة مثل التابير) والproboscideansوغيرهم))من نهاية عصر البليوسين من حوالي 60 مليون سنة تقريباً.
الخصائص التي تربط الSinonyx بالحيتان والتي بالتالي توضح قرابتهم ..
1- فم ممدودelongated muzzles 2- jugular foramen (فتحة في قاعدة الجمجمة) واسعة 3- basicranium (التكوين العظمي الداخلي للجمجمة) قصير (Zhou and others 1995).
عدد الأسنان كان للثدييات البدائية (44)؛ الأسنان متمايزة كما الأسنان الheterodontللثدييات الحالية. الضروس كانت أسنان قص ضيقة جداً خصوصاً بالفك السفلي لكن بها عدة نتوءات cusps. الفم الممدود في الأغلب لأستخدامه في صيد الأسماك؛ كما الحيتان والدلافين التي تصطاد الأسماك بمساعدة فمها الممدود للأمام.
هذه الخواص هي غير مماثلة بشكل كامل للmesonychids، موضحة أن الSinonyx كان منذ وقتها يطور خصائص تكيفية التي ستكون لاحقاً أساساً لحياة الحيتان التخصصية في الماء.
Pakicetus
الحفرية الثانية في ترتيبنا هي Pakicetus وهو أول حوت بدائي- archaeocete - معروف حتي الآن. هي من بدايات عصر الأيوسين من 52 مليون سنة مضت(Gingerich and others 1983).
وعلي الرغم من إنها معروفة فقط من بقايا أجزاء للجمجمة (Pakicetusinachus)، فهذه البقايا تشخص وضعها بشكل صريح كنوع وسط بين ال Sinonyxوالحيتان؛ وتحديداً من ناحية الأسنان.
الضروس العلوية والسفلية والتي تبقي لها عدة نتوءات cusps وبالتالي تبقي مشابهة للحفرية السابقة Sinonyx؛ لكن الطواحن premolars أصبحها كتشكيل أكثر بساطة كأسنان مثلثة بنتوء واحد ومسننة من حرفها الأمامي والخلفي. أسنان الحيتان التي تطورت في مرحلة لاحقة ظهرت فيها فيها شكل أكثر تبسيطاً أيضاً للمثلثات المسننة كمثل الموجودة في أسماك القرش آكلة اللحوم؛ موضحة أن أسنان الPakicetus تكيفت لصيد الأسماك.
Pakicetus لم يسمع بشكل جيد تحت الماء. جمجمتها لم تحوي tympanic bullae -فقاعة طبلية- كثيفة أو تجاويف تعزل الauditory area -الباحة السمعية- اليسري عن اليمني، وهذه خصائص ظهرت في الحيتان والحفريات التالية بعدها مكنتهم من القدرة علي السماع وتمييز إتجاهات الموجات الصوتية تحت الماء وتمنع إنتقال الأصوات خلال الجمجمة (Gingerich and others 1983).
في الأغلب Pakicetus لم تكن لديه القدرة علي الغوص أو الحياة تحت الماء بأي عمق كان. هذه التقييم للحفرية متوافق تماماً مع الأدلة الجيوكيميائية والبيئية التي سنتحدث عنها بعد قليل. في النهاية الPakicetus كان كائن أرضي أكثر منه مائي. لكن يبقي شكل الجمجمة الممدود -elongated muzzlesوبشكل أظهرمن الSinonyx - قطعاً من خواص الحيتان (cetacean) وأسنانها كما شرحنا توضح بشكل ظاهر حالة إنتقالية بين الأقدم والأحدث –Sinonyx&Modern Whales- في طريق تطور الحيتان الذي تطرحه نظرية التطور وتتوافق معه.
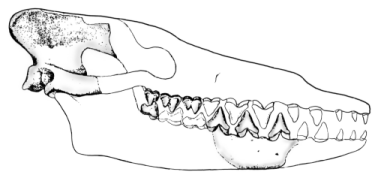
إعادة تشكيل لجمجمة (Pakicetusinachus) قام بها Philip D. Gingerich وآخرين عام 1983
وهنا تجدر الإشارة لأحد أهم إعتراضات معارضي التطور للحفرية السابقة وهي أن الحفرية مجرد أجزاء من الجمجمة لا تصلح لإثبات حقيقة أن الPakicetusجزء من حلقة التطور للحوت من الثدييات الأرضية ونقتبس هنا ما نشرته مؤسسة Answers in Genesis (AiG) التي تدعم منشورات دورية معارضة ورافضة للتطور وذلك عام 1996 " Pakicetus consists of only a few skull fragments.(!) " أي أن حفرية Pakicetus هي عبارة عن فقط فتات من جمجمة !
هذا كان صحيح في حالة Pakicetusinachus التي إكتشفت عام 1981 بواسطة P. D. Gingerich and D. E. Russell، علي الرغم من أن البقايا المكتشفة تكشف بوضوح مكانها كحلقة وسيطة في سلسلة تطور الحوت؛ لكن تعدل الوضع بعد ذلك في عام 2001 حينما أكتشفت حفرية شبه كاملة للPakicetusسميت بالPakicetusattocki من فريق من العلماء من جامعات أوهايو وأريزونا وهاورد وجاءت الحفرية الجديدة لتأكد الإستنتجات السابقة من جمجمة Pakicetusinachus وخصوصاُ أن الPakicetus هو كائن أرضي أكثر منه مائي.
للإطلاع أكثر ..
http://www.nature.com/nature/journal/v413/n6853/full/413277a0.html
http://www.faculty.virginia.edu/bio202/202-2002/Lectures%2020202/thesissen%20et%20al%202001.pdf

Pakicetusattocki(متحف أونتاريو الملكي، كندا)
Ambulocetus

Ambulocetusnatans (متحف التاريخ الطبيعي بيزا، إيطاليا)
في نفس المنطقة التي أكتشف فيها الPakicetus في باكستان، لكن في ترسبات أعلي بحوالي 120 متر. قام Thewissen وآخرين في 1992 بإكتشاف Ambulocetusnatans(من منتصف لبداية عصر الأيوسين من حوالي 50 مليون سنة). حفرية Ambulocetus كانت إكتشاف رائع فكانت بوضوح cetacean–حوت- مع أرجل وهيكل يسمح لها بدرجة من القدرة علي الحركة علي الأرض.
ما إستنتجناه عن قدرة الAmbulocetusعلي المشي بأطرافها الخلفية (hind limbs) يدعمه كونها كبيرة، مع عظمة فخذ سميكة (stout femur). وعلي الرغم من ذلك وبسبب أن عظمة الفخذ لم تملك نقاط ربط عريضة والضرورية للعضلات للمشي، فلا يمكنها بذلك أن تكون كائن متحرك بشكل كفؤ علي الأرض وعلي الأرجح كانت نمط حركتها شبيه بكائنات أسود البحر الحديثة عن طريق دوران قدمها الخلفية للأمام ثم الحركة المتهادية علي الأرض بدعم من قدمها الأمامية والإنثناء المرن لعمودها الفقري.
هذا عن الأطراف الخلفية، أما عن الأطراف الأمامية (forelimbs) فهي أيضاً تمثل حلقة وسط بين الكائنات الأرضية والمائية من حيث التركيب أو الوظيفة. عظمتي الulnaوradiusكانتا قويتين بحيث كانت قادرة علي حمل وزن الحيوان علي الأرض؛ الكوع قوي تركيبياً لكنه فقط يميل في الحركة للخلف فقط وهذه ميزة تطورية لهذا النوع ساعدته علي دفع الساعد (forearm) بقوة للخلف للسباحة تحت الماء. لكنه وعلي الرغم من ذلك كان له رسغ (wrist) مرن وهذا خاصية ليست موجودة في الحيتان الحديثة كما ذكرنا.
ومما هو واضح أيضاُ من تشريح العمود الفقري للAmbulocetusأنه كانت له القدرة علي العوم بحركته المتمايلة للأعلي وللأسفل مع دفع من قدمه الخلفية في إتجاه الخلف.
ويجدر هنا الإشارة إلي أن أصابع قدمه الخلفية تنتهي بحوافر (hooves) وهذا بالطبع له علاقة بأصول هذا الكائن مع أسلافه ذات الحوافر (ungulates).
أما جمجمة الAmbulocetus فكانت أقرب كثيراً للحيتان (cetacean) (Novacek 1994). بحيث أن لها long muzzle و أسنانها كانت شبيهة جداً بنوع حديث سيأتي بعدها وأقرب للحيتان وهو (archaeocetes)، عظم وجنة مصغر (zygomatic arch) وأيضاً فقاعة طبلية (tympanic bullae) –التي تحمل طبلية الأذن- مربوطة بشكل ضعيف مع الجمجمة. ويلاحظ مع تقارب خصائص جمجمة الAmbulocetus مع الحيتان الحديثة إلا أنه مازال يفتقد لفتحة النفخ (blowhole).
علي العموم فأفضل توصيف للAmbulocetus هو كائن بر-مائي بحجم أسد البحر آكل للأسماك لكنه لم يكن فاقد للكلية لأصوله الأرضية سواء تشريحياً أو كنمط للحياة.
Rodhocetus
من منتصف عصر الأيوسين (46.7 مليون سنة)، أخذ Rodhocetus كل التغيرات لسابقيه ولمرحلة أكثر تقدماً في التكيف مع الحياة البحرية، لكن مع الإحتفاظ بخواص بدائية للحياة علي الأرض (Gingerich and others 1994).
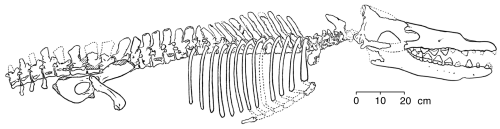
Rodhocetuskasrani, Gingrich and others (1994)
هي أول archaeocete (حيتان بدائية) بفقرات صدرية (thoracic)، أسفل الظهر (lumbar) وعجزية (sacral) ظلت محفوظة.
فقرات أسفل الظهر لها أشواك عصبية (neural spines) أكثر من سابقاتها من الحيتان الأقدم. وحجم هذه الإمتدادات في قمة الفقرات عند ربط العضلات توضح أن الRodhocetusله ذيل متطور وقوي للسباحة أكثر من سابقيه.
أيضاً الفقرات العجزية الأربعة الكبيرة غير مدموجة في بعضها، مما يعطي للعمود الفقري مرونة كبيرة وسمحت أكثر فأكثر لدفع قوي أثناء السباحة.
وكان –علي الأغلب- الRodhocetus له ذيل fluke مثل الحيتان الحديثة ومع أن هذه الصفة لم تبق في السجل الحفري ولم تحفظ لهذا النوع لكن لها خصائص دالة عليها مثل: فقرات عنقية قصيرة، فقرات ذيل ثقيلة وقوية، أشواك ظهرية (dorsal spines) خلفية كبيرة ملائمة لذيل كبير ولأربطة العضلات الأخري. هذه الخصائص هي نفسها المرتبطة بالحيتان الحديثة مع تطور وإستخدام الذيل (tail flukes).
في كل الأحوال الRodhocetus كان سباح ماهر بذيله وبذلك هو يعتبر أقدم حفرية للحيتان تظهر هذا الأسلوب في العوم.
حوض الRodhocetusكان أصغر من أسلافه لكنه كان مازال وقتها متصلاً بالفقرات العجزية (sacral vertebrae) مما يعني أنه كان في هذه المرحلة مازالت له القدرة علي المشي علي الأرض بشكل محدود. مع ذلك فقصر الحوض مقارنة بالmesonychidsيجعله أقل قدرة وإمكانية من التكوين العضلي علي دفع نفسه للأمام أثناء الحركة الأرضية؛ عظم الفحذ (femur) أقصر بمقدار الثلث من الAmbulocetus ولذلك الRodhocetus لم يكن له القدرة علي الإستمرار في الحركة الأرضية كوقت وسرعة وإمكانية (Gingerich and others 1994).
جمجمة الRodhocetusكانت كبيرة مقارنة بباقي الهيكل العظمي، الpremaxillaeوdentaries(تكوينات عظمية في الفكين العلوي والسفلي) ممتدين للأمام بشكل أكبر من أسلافه مما يزيد من إمتداد الجمجمة بشكل بارز للأمام ويجعلها أقرب وأقرب للحيتان -more cetacea- عن أسلافه.
الطواحن –molars- لديها crownsأعلي من من الحيتان المبكرة وأكثر تبسيطاً. الطواحن السفلية أطول منها لعرضها. وللمرة الأولي إنتقلت فتحات التنفس للخلف بطول الsnout-الإنف- وتمركزت فوق الناب -the canine teeth- كدليل علي تطور الblowholeعند الRodhocetus مثل الحيتان الحديثة.
الفقاعات السمعيةThe auditory bullae كبيرة وتتكون من عظام سميكة وهي خصائص مميزة وفريدة للحيتان، لكن لا يبدو أن الجمجة تحتوي علي جيوب أنفية –sinuses- مماثلة للحيتان التالية والأحدث، مما يثير التساؤل حول قدرة الRodhocetusعلي تحديد الإتجاهات عند السماع تحت الماء.
وبشكل عام، الRodhocetusأظهر خواص مطورة عن الحيتان المتقدمة بفقرات صدرية عميقة ورفيعة، رأس أطول، مرونة أعلي للفقرات وتكوين عظمي خاص بالذيل أكبر. فمن ذلك نجد مرونة وقوة أكبر في الظهر ومنطقة الذيل والعكس في حالة الأطراف مما يعطي تأكيد علي كون الRodhocetus كان سباح ماهر بإستخدام الذيل مع قدرة أقل للحركة علي الأرض.
Basilosaurus
.jpg)
Basilosauruscetoides(المتحف الوطني للتاريخ الطبيعي، واشنطن)
حفرية ال Basilosaurusالشهيرة تمثل الخطوة التطورية التالية في طريقة تطور الحيتان.عاشت في نهاية عصر الأيوسين ونهاية منتصف الأيوسين (35-45 مليون سنة). (Gingerich 1994)
الBasilosaurus هو حيوان طويل، رفيع وملتوي كان في الأصل يعتقد أنه بقايا لثعبان بحر (ومنه أتي أسم الحفرية التي تعني king lizard). طوله الهائل (يصل ل15 متر) يبدو أنه نتيجة لخاصية فريدة بين الحيتان؛ فقراتها ال67 طويلة جداً بالمقارنة بالحيتان الأخري في هذا الوقت وحتي للحيتان الحديثة ولذلك من المحتمل أن يمثل هذا النوع نوع من التخصص (specialization) التي تفصلها عن الخط الذي سار عليه تطور الحيتان الحديثة.
ما يجعل الBasilosaurus علي وجه الخصوص حوت مثير للإهتمام والبحث، هي خصوصية تشريح أطرافه الخلفية (hind limbs) (Gingerich and others 1990).فهي بالكاد عندها تركيب حوضي كاملcomplete) pelvic) ومجموعة من عظام الأطراف الخلفية.
الأطراف كانت صغيرة جداً لأي دفع فعال، أقل من 60 سم في مقابل طول يصل ل15 متر والحوض كان منفصل بشكل كامل عن العمود الفقري مما يجعل إحتمال الكائن لوزنه مستحيلاً. ومع وجود أقدامه خارج الجسم، نري وبشكل واضح حلقة وسط في طريق تطور الحوت من الحياة الأرضية للبحرية من الناحية التشريحية.
وعلي الرغم من عدم وجود ذيل الحوت المميز (tail fluke) –ويرجع ذلك لأن ذيل الحوت لا يحوي عظام وبالتالي ليس من المحتمل أن يتحجر- قام Gingerich وآخرين في 1990 بملاحظة أن ال Basilosaurus له عمود فقري يتشارك في خصائص مع الحيتان التي تملك tail fluke.
الذيل والفقرات العنقية أقصر من الأجزاء المكونة لمنطقة الصدر وخلف الظهر، ومنها أخذ الباحثون تناسب الفقرات كدليل علي إحتمالية إمتلاك الBasilosaurus ذيل الحوت المميز.
دليل آخر علي أن الBasilosaurus عاش أكثر حياته تحت الماء يأتي من تغير آخر مهم في الجمجمة. الحيوان كان يملك فتحة أنف واحدة كبيرة إنتقلت مسافة قصيرة للخلف لنقطة تناظر الثلث الخلفي من صف الأسنان.
ويعد إنتقال الأنف (snout) من مقدمة الجمجمة لوضع أقرب لمؤخرة الرأس هي خاصية مميزة فقط للثدييات التي تعييش في بيئة بحرية أو مائية.
Dorudon
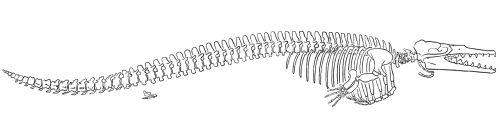
Gingrich and Uhen (1996) published this reconstruction of the skeleton of
Dorudonatrox (redrawn for RNCSE by Janet Dreyer).
Dorudonatrox (redrawn for RNCSE by Janet Dreyer).
الDorudon كان معاصر للBasilosaurus في نهاية عصر الأيوسين (منذ حوالي 40 مليون سنة) ومن الأرجح أنه يمثل المجموعة التي تعد السلف للحيتان الحديثة أكثر من الBasilosaurus(Gingerich 1994).
وكان الDorudonيفتقر للتكوين الفقاري الطويل الذي ميز الBasilosaurus (فقط حوالي 4-5 متر في الطول).و ترتيب الأسنان للDorudonمشابه للBasilosaurus، ال cranium-الجمجمة وتحديداً الجزء الذي يغلف المخ- ومقارنة بجمجمة الBasilosaurusوالحيتان المتقدمة كانت أكثرتقوساً (vaulted)(Kellogg 1936).
الDorudonلم يكن أيضاً يملك تشريح للجمجمة تظهر وجود جهاز أو تركيب قادر علي تحديد إتجاهات الأشياء تحت الماء عند إنعكاس الأصوات المرسلة (echolocation) (Barnes 1984).
ومن السابق نستخلص أن الBasilosaurus والDorudon يعتبروا كائنات مائية بشكل صريح - الDorudon مثل الBasilosaurus يملك أطراف خلفية صغيرة جداً تظهر بشكل طفيف تظهر خارج جسمه- وهم بالتالي لم يعودوا مرتبطين بأي شكل بالحياة الأرضية؛ في الحقيقة لم يكن بإمكانهم الحركة علي الأرض علي الإطلاق. حجمهم وإفتقارهم للأطراف الضرورية لدعم وزنهم علي الأطراف جعلتهم كما قلنا بالضرورة كائنات مائية، وهو توجه تم تفصيله وتأكيده في تصنيف الحيتان (whale taxa) لاحقاً.
بشكل واضح مما سبق، وحتي لو أكتفينا بالدليل الأول من السجلات الحفرية (paleontological evidence) سنجد إدعاءات الخلقيين ومعارضي التطور في أنه " لا حلقات وسط من الحفريات " بين الثدييات الأرضية والحيتان هي خاطئة !.
فعلي العكس تماماً من ناحية الحيتان–كما شرحنا- سنجد عدد من الحفريات المرتبة مورفولوجياً وزمنياً بشكل متناسق يأكد حقيقة تطور الحوت من الثدييات الأرضية.
وكملخص لما ذكرنا في الدليل الحفري، سنجد تغيرات متناسقة توضح مجموعة من الخصائص تظهر حالات متتابعة من التكيفات من إتجاه الحياة الأرضية في إتجاه الحياة المائية كلما توجهنا من السلف الأبعد للأنواع الأحدث. هذه التغيرات تظهر في شكل الجمجمة، شكل الأسنان، وضع ومكان الأنف، حجم وتركيب الأطراف الأمامية والخلفية، حجم وشكل الذيل و تركيب الأذن الوسطي التي ترتبط بسماع الإتجاهات تحت الماء.
أخيراً لا ننسي تأكيد أن الدليل الحفري يسجل تاريخ من التكيف المتزايد في إتجاه الحياة تحت الماء، ليس فقط أي شكل للحياة تحت الماء، بل تحديداً لحياة تعيشها الحيتان الحالية.
2- الدليل التشكيلي (Morphological evidence)
دراسة الخصائص المورفولوجية التي تشترك فيها حفريات الحيتان -التي سبق شرحها في الأعلي- والثدييات ذوات الحوافر (ungulates) المعاصرة يظهر وجود السلف المشترك بينهما بشكل أكثر وضوحاً.
- فعلي سبيل المثال تشريح قدم الBasilosaurus يربط الحيتان مع ذوات الحوافر من النوع (artiodactyls) (Gingerich and others 1990)، فمحور تماثل القدم في حفريات الحيتان يقع بين الأصبعين الثالث والرابع فيما يعرف هذا التنظيم ب(paraxonic) وهذه خاصية مميزة للحيتان وذوات الحوافر artiodactyls&condylarths ونادرة الوجود في مجموعات حية أخري (Wyss 1990).
- مثال آخر يظهر ال incus(سندان الأذن الوسطي) في حفرية الPakicetus-المحفوظ علي الأقل في عينة واحدة- من الناحية المورفولوجية وسط في كل الخصائص بين ال incusفي الحيتان الحديثة ومثيلاتها في ذوات الحوافر artiodactyls الحديثة (Thewissen and Hussain 1993).
3- الدليل الجزيئي (Molecular biological evidence)
الفرضية التي تطرحها نظرية التطور عن الحيتان وإنحدارها من أسلاف ثدييات أرضية يتبعه بالتالي وجود تشابهات علي المستوي الجزيئي والحيوي بين الحيتان والثدييات الأرضية الأقرب لها بشكل يدعم حقيقة تطورهما من سلف مشترك واحد.
وينتج عن هذا وبشكل عام أنه كلما تقاربت أنواع أو مجموعات من الكائنات زادت التشابهات بينهم علي المستوي الجزيئي والشكلي والسلوكي إلخ والعكس صحيح؛ وهذا لا يكون له أي معني فقط إلا بتفسيره عن طريق حقيقة التطور والسلف المشترك بين الأنواع فلو كان كل كائن خُلق بمفرده بمعزل عن غيره لما كان لتوزيع درجة التطابقات علي المستوي الجزيئي بين الكائنات قرباً وبعداً له أي معني أو تفسير علمي.
الدراسات الجزيئية لجودمان وآخرين (1985) توضح أن الحيتان هي أقرب لذوات الحوافر ungulates من كل الثدييات الأخري (تنبغي الأشارة هنا إلي أن الأبحاث الأحدث وضعت كائن ال hippo أقرب الأنواع الحية للحيتان من الناحية الجزيئية وهذا الإكتشاف أعاد ترتيب التصور لمسار تطور الحوت وإن كان لم يأثرعلي حقيقة أن أصل الحوت يرجع كما نشرح لثدييات أرضية أو أن الحيتان لها أسلاف مشتركة مع ذوات الحفرياتartiodactyls))وسنفصل هذه النقطة في نهاية المقال).
هذه الدراسات إختبرت ال myoglobin, lens alpha-crystallin A, and cytochrome cفي دراسة من 46 نوع مختلف من الثدييات. مياموتو وجودمان (1986) لاحقاً قاموا بتوسيع عدد ال protein sequences(ترتيب البروتين من الناحية الجزيئية) بإضافة alpha- and beta- hemoglobins and ribonuclease؛ أيضاً زادوا عدد الثدييات في الدراسة ل72. وفي كل الأحوال كانت النتائج متطابقة؛ الحيتان وبشكل واضح أقرب لذوات الحوافر(ungulates).
ولا بأس من الإشارة إلي دراسات جزيئية أخري شملت دراسة جينات، بروتينات وإنزيمات بواسطة
Irwin and others (1991), Irwin and Arnason (1994), Milinkovitch (1992), Graur and Higgins (1994), Gatesy and others (1996), and Shimamura and others (1997)وأيضاً أوضحت نتائجها أن الحيتان هي كائنات مقربة من ذوات الحوافر (artiodactyls) مع ملاحظة وجود بعض الإختلاف في تفاصيل هذه الدراسات.
مع النتائج السابقة التي توضح حقيقة تقارب الحيتان مع ذوات الحوافر (artiodactyls)، فالدراسات الجزيئية تأكد الفرضيات التي تطرحها نظرية التطور وبشكل صريح، فهذا النمط من التشابهات علي المستوي الجزيئي والحيوي يجب أن يظهر فقط في حالة أن الحيتان وذوات الحوافر (ungulates) وتحديداً ال Artiodactyls يتشاركون في سلف مشترك واحد.
4- الدليل الأثري (Vestigial evidence)
الخصائص ال vestigialللحيتان تأكد لنا علي شيئين.أن الحيتان مثل عديد الكائنات الأخري لها خصائص لا تحمل أي معني أو منطق لو أفترضنا أنها كانت نتيجة عملية تصميم؛ فليس لها أي وظائف أساسية كمثيلاتها في الأنواع الأخري ومستهلكة للطاقة بل قد تكون مضرة بالكائن. وتأكد لنا أيضاً أن الحيتان تبقي فيها أثار من ماضيها التطوري وهو في هذه الحالة سلف أرضي.
فالحيتان الحديثة في الغالب تحتفظ ببقايا من عظام الحوض وعظمتي ال femoraو tibiaeأقرب لشكل عمود ومدمج في التكوين العضلي لأجسامهم. هذه العظام تظهر بشكل أوضح في الأنواع المتقدمة وتتقلص في الأنواع الأحدث، فعلي سبيل المثال الBasilosaurus له حجم أكبر من بقايا الحوض وعظام الأطراف الخلفية من الحيتان الحديثة وبالتأكيد أقل بكثير من الأنواع التي سبقته –وتم شرحها هنا- وعاشت حياة أرضية أكثر منها مائية.
الحيتان أيضاً تبقي عندها عدد من الخصائص ال vestigialمن أسلافها الأرضية في أجهزتها الحسية، نذكر منها:
- الحيتان الحديثة لديها أعصاب للشم وهي فقط vestigial وتوضح بجلاء إرتباط الحوت بأصوله الأرضية.
- أيضاً في الحيتان الحديثة الفتحة الخارجية للقناة السمعية (the auditory meatus) مغلقة. في العديد منهم هي في حجم 1مليمتر تقريباً وفي الأغلب هي منضغطة في منتصف المسافة.
- كل الحيتان تملك عدد من العضلات الصغيرة يفترض إحتياجها في عملية تحريك الأذن الخارجية –التي لا يملكها الحوت الحديث- وإحتاجها سلفه الأرضي في تحريك أذنه (directional hearing) وهذا سلوك حصري للحيوانات الأرضية ومرة أخري دليل جديد علي إنحدار الحوت من أصول أرضية.
- الحجاب الحاجز في الحيتان هو vestigial ويملك عضلات ضعيفة جداً. الحيتان تستخدم الحركة الخارجية للضلوع لملء الرئة بالهواء.
- الحجاب الحاجز في الحيتان هو vestigial ويملك عضلات ضعيفة جداً. الحيتان تستخدم الحركة الخارجية للضلوع لملء الرئة بالهواء.
أخيراً، رصد عدد من الحالات لأجنة الحيتان بواسطة Gould (1983) يظهر فيها أطراف خلفية (hind limbs) بارزة. وبالمثل رصدت أجنة دلافين ب finsدقيقة خاصة بالحوض، علي الرغم من أنهم لم يكونوا مدعومين بعظام أطراف (limb bones) كما في حالة أجنة الحيتان.
وفي بعض الحيتان مثل الحوت الأبيض (beluga)، نجد أجزاء بدائية غير كاملة من الجزء الخارجي للأذن؛ وهي خاصية ليس لها أي هدف ظاهر لحيوان لا يملك أذن خارجية وبالعكس قد تسبب ضعف في كفاءة السباحة للحيوان بزيادة مقاومة الماء hydrodynamic drag)) لحركته.
وعلي الرغم من أن قائمة الخصائص الأثرية التي شرحناها ليست شاملة أو مفصلة، لكنها تبقي مع ذلك واضحة وصريحة أن الحيتان عندها غني من الخصائص ال vestigial من أسلافها الأرضية.

هيكل الDorudon، ويلاحظ وجود ال hind limbsمنفصلة عن الهيكل الفقاري له. (2)

الحوت القاتل (Killer whale, Orcinus orca) ، ويلاحظ وجود ال hind limbsمنفصلة عن الهيكل الفقاري له. (3)
5- الدليل الرجوعي (Atavistic evidence)
وهو واحد من الأدلة المهمة التي تؤكد حقيقة تطور الحوت من الثدييات الأرضية، ففي بعض الحالات يحتفظ الحوت بآثار –أجزاء من الجسم- لتكون جزء منه بعد الولادة ليست من التركيب أو التشريح الأصلي له، في نفس الوقت التي تعتبر جزء أساسي لأسلافه الأرضية التي تفترضهم نظرية التطور.
مثال علي ذلك:

الأسهم الصفراء تظهر أطراف خلفية كاملة كشكل من أشكال الرجوعية Atavism لدلفين وجد في المياه الساحلية اليابانية بتاريخ 28 أكتوبر 2006 (4)
6- الدليل الجنيني (Embryological evidenc)
مثل الخصائص ال vestigial، الخصائص الجنينية (embryological) تُأكد لنا علي نقطتين. الأول أن جنين الحوت يطور مجموعة من الخصائص التي يفقدها لاحقاً قبل وصوله لشكله النهائي قبل الولادة. وهنا علي معارضي التطور تفسير مثل هذه العملية اللامعقولة في أن تبني تركيب عضوي معين قبل أن تفقده لاحقاً دون سبب له فائدة للنوع ؟!
داروين في العام (1859) سأل نفس السؤال. أليس من المنطق أن يتشكل الجنين بشكله النهائي أسرع وبشكل مباشر؟ لاتبدو علي الأطلاق هذه عملية ذات معني لو كانت نتيجة لعملية تصميم محددة من قوة خارجية أن ترسل الجنين في هذا المسار كثير التغيرات بدون حاجة !، لكن التطور كعملية تتطلب بناء خصائص جديدة علي أساس من خصائص أسلاف النوع السابقة تعدلها أو تزيلها لاحقاً وهذه النقطة هي حجر زاوية في حقيقة التطور فلا إنفصال كامل بين النوع وأصوله وهذا ما تعاكسه نظرية الخلق المنفرد للأنواع التي تفترض أن كل الكائنات مخلوقة بشكل منفصل تماماً عن غيرها.
النقطة الثانية، أن دراسة جنين الحوت بالتفصيل تعطينا وبشكل واضح دلائل علي أصوله الأرضية. الجنين ليس أقل من الحوت البالغ في إحتواءه علي أثار ظاهرة وغزيرة من الخصائص الأرضية التي فقدت وظيفتها مع تكيف الحوت مع الحياة المائية.
حيتان عديدة وهي مازالت جنين في الرحم تبدأ في نمو شعر علي جسدها. وعلي الرغم من ذلك فالحيتان الحديثة لا تحتوي علي أي شعر علي أجسادها بعد الولادة، ما عدا بعض الشعيرات حول ال blowholesتستخدم للإستشعار في بعض الأنواع. حقيقة أن الحياة تمتلك جينات لأنتاج شعر لأجسامها يظهر أن أسلافهم كانت تغطي أجسادهم الشعر. ببساطة كانت ثدييات أرضية عادية.
أيضاً في العديد من أجنة الحيتان، نجد بروز لأطراف خلفية (hind limb buds) قبل أن تختفي في مراحل تطور الجنين مع الوقت.
أيضاً نري بعض أجزاء بدائية للجزء الخارجي من الأذن (are rudimentary ear pinnae ) وأيضاً تختفي مع تطور الجنين وقبل الولادة (ماعدا في بعض الحالات النادرة كأحد الخصائص الرجعية atavism والتي هي الأخري من أهم الأدلة علي حدوث التطور كما شرحنا في الأعلي). وفي بعض الحيتان فصوص الشم للمخ نجدها أيضاً وفقط في الجنين قبل أن تختفي مع مراحل تطوره.
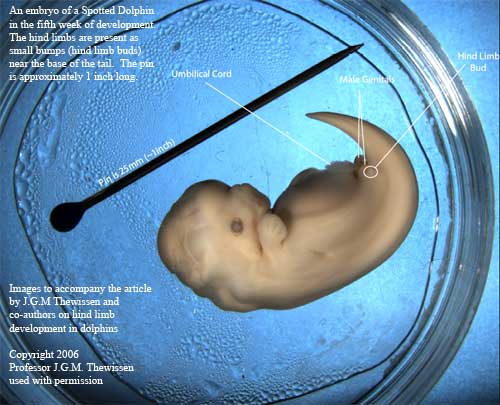
صورة لجنين دولفين في أسبوعه الخامس. تظهر فيها بوضوح أطراف خلفية (hind limbs) بالقرب من قاعدة الذيل وهذه الأطراف طولها تقريباً 1 بوصة (2006, Dr. J.G.M. Thewissen)
أيضاً فتحتي الأنف (nostrils) هما في وضعهم الطبيعي للثدييات في مقدمة الأنف (snout) في بداية تكوين الجنين لكن لاحقاً ومع نموه، الفتحات تنتقل للخلف لمكانهم الأخير في قمة الرأس ليشكلوا الblowhole(s).
حتي عملية تطور أنواع الحيتان من بينها يمكن فهمها عن طريق دراسة أجنتهم (embryology). نحن الآن نعرف أن حيتان ال baleenتطوروا من الحيتان ذات الأسنان (toothed)، فبعض أجنة حيتان ال baleenتبدأ في نمو الأسنان بجانب شعر الجسم قبل أن يختفي كل هذا لاحقاً مع تطور الجنين قبل الولادة. ولأن وجود أسنان عند الجنين في الرحم ليس له فائدة للحيوان –ولم يطرح حتي الآن معارض التطور فائدة يمكن الإستناد عليها علمياً- ففقط إرجاع وجودها للعوامل الوراثية التي تربط الحوت بأصوله الأرضية هي ما تعطي لهذا الظاهرة العجيبة معني وتفسير علمي رصين؛ فلا يوجد علي الإطلاق لو أفترضنا فرضية الخلق المنفرد للأنواع أي معني أو منطق أن نعطي لأجنة الbaleen أسنان ؟!
في الأخير لدينا من جديد مجال علمي مستقل يعطينا نتائج وحقائق متناسقة ومترابطة مع كل ما شرحناه –وسنشرحه- من المجالات العلمية المختلفة وكلها تصب في خانة واحدة فقط: الحيتان تمتلك خصائص ظاهرة تربطها بأصولها وأسلافها من الثدييات الأرضية وتحديداً ذوات الحوافر.
7- الدليل الجيو كيميائي (Geochemical evidence)
الحيتان المتقدمة عاشت في بيئة عذبة (freshwater)، لكن أسلاف الحيتان الحديثة بعد ذلك إنتقلت للحياة في بيئة مالحة (saltwater) –كما هو الحال الآن- وبالتالي كان من الواجب عليها التكيف لشرب الماء المالح.
وبما أن المياه العذبة والمالحة تختلفان في نسبة نظائر الأكسجين (isotopic ratios of oxygen) ويمكننا التنبؤ بهذا الإنتقال من بقايا حفريات الحيتان وتحديداً الأسنان. وهذا ما رأيناه فحفريات الأسنان من الحيتان المتقدمة لديها نسبة أكسجين ثقيل لخفيف أقل مما يؤكد حقيقة أن هذه الحيوانات كانت تشرب من المياه العذبة (Thewissen and others 1996).
فيما نري أن حفريات الأسنان للحيتان المتأخرة نسبة أعلي من الأكسجين الثقيل للخفيف، ما يوضح أنهم كانوا من شاربي المياه المالحة. كل هذا وبدون شك يدعم الإطار التي تفرضه نظرية التطور والذي ناقشناه حتي الآن: أسلاف الحيتان الحديثة تكيفت من سلوكيات الحياة الأرضية إلي الماء المالح مروراً بعادات الحياة في البيئة العذبة.
8- الدليل الحفري-بيئي (Paleoenvironmental evidence)
في هذا الدليل نطرح منطقية تسلسل حفريات الحيتان والتغيرات فيها بأنها يجب أن تكون أيضاً متناسقة مع التغيرات في حفريات الكائنات الأخري في نفس البيئة والتوقيت؛ فحفريات الكائنات الأخري التي وجدت بجانب حفريات الحيتان يجب أن توضح بجلاء الوضع للبيئة التي عاشت فيها الحيتان وقتها. ولا ننسي أيضاً أن يتوافق ويتناسق هذا الدليل مع كل الأدلة السابقة التي سبق تناولها. وبالتالي هنا يجب أن نجد دليل علي عدة تحولات في البيئة التي عاشت فيها الحيتان من أرضية وحتي نصل إلي بيئة بحرية بالكامل وعلي هذا الترتيب نجد حفريات الحوت.
مورفولوجيا الSinonyx تظهر بوضوح أنه كان كائن أرضي، ولذلك لا تعد هنا مفاجأة أن نجد حفرياته فقط مع كائنات أخري برية. الPakicetus ورغم أنه كان أرضي أكثر منه مائي لكن من الممكن طبقاً لتشريحه أن كان يتواجد لفترات طويلة في الماء بحثاً عن الغذاء، وعلي الرغم من أننا وجدنا حفريات لثدييات أرضية بجانب حفرياته من القوارض والوطاويط وذوات حوافر وحتي رئيسيات (Gingerich and others 1983)، فيوجد أيضاً حيوانات مائية كالقواقع، أسماك، سلاحف وحتي تماسيح من الرتبة crocodilian.
أيضاً الترسبات التي تواجدت بها حفرية الPakicetus تظهر بجلاء حدوث تدفقstreaming or) flowing)، وهذا فقط مصاحب للتربة المحملة بالماء.
الدليل الpaleoenvironmentalيظهر لنا بوضوح أن الPakicetus عاش في بيئية أرضية منخفضة ورطبة مع تواجد في مناطق الماء العذب بحثاً عن الغذاء. أيضاً وبشكل مثير للإهتمام نجد أسنانه الدائمة واللبنية في هذه الترسبات بنفس قدر التواتر تقريباً مما يدعم فكرة أن الPakicetus يقوم بعملية الولادة علي الأرض.
الترسبات التي وجدت فيها الAmbulocetusوجدت تحتوي علي leaf impressions بالإضافة إلي حفريات للحلزون والرخويات البحرية الأخري (marine mollusks). هذه التركيبة تظهر أن الAmbulocetusوجدت فيما يمكن أن يكون في السابق بحر ضحل (shallow sea).
الRodhocetus وجدت في Shales (صخور رسوبية طينية) خضراءمترسبة في عمق ال neritic zoneتقريباً في حدود ال200 متر تحت الماء (بشكل محازي للجزء الخارجي من ال.(continental shelfوهذه ال green shales هي فقط تتواجد في المياه العميقة منخفضة نسبة الأكسجين، وبالتالي نستنتج أن الRodhocetus عاش في أعماق أكبر من كل الحيتان المتقدمة عنه. وأيضاً حقيقة أن حفرية الRodhocetus وجدت مصاحبة لكائنات المنخربات البلانكتونية (planktonic foraminiferans) وغيرها من الحفريات المجهرية تأكد مع إستنتجناه علي درجة عمق المياه التي عاش فيها الRodhocetus .
الBasilosaurus والDorudonhaveوجدوا في ترسبات مختلفة (Kellogg 1936)، مما يظهر قدرة النوعين علي الحياة في المياة العميقة وحتي الضحلة.
أخيراً مما شرحناه -ولو بإختصار- من حيث الدليل الحفري-بيئي (paleoenvironmental)، يمكننا القول أن الحيتان وهم في مسارهم التطوري كانوا في إتجاه ونمط حياة متزايد في إتجاه المياه العميقة مع إنفصال وإنسلاخ تدريجي عن بيئتهم الأرضية وحتي القريبة من الشواطئ وهو ما يتوافق مع كل الأدلة العلمية التي تم شرحها حتي الآن.
9- الدليل الحفري-حيوي-جغرافي (Paleobiogeographic evidence)
الدليل الجغرافي هو أيضاً متناسق مع نمط توزيع الحيتان المتقدمة ثم إتساع أماكن التواجد الجغرافي لها بعد ذلك. هنا من المنطقي أن نتوقع توزيع للأنواع الأرضية في مساحات جغرافية محددة علي عكس الأنواع المائية، التي يعد كامل المحيط بالنسبة لها هو المدي الجغرافي لإنتشارها. فمدي توزيع الSinonyx فقط في أسيا الوسطي. عينات الPakicetus وجدت فقط في باكستان؛ الAmbulocetusوالRodhocetus وجدوا أيضاً في مناطق محددة. وعلي العكس فالBasilosaurus والDorudonالحفريات التي تمثل الحيتان المتأخرة والتي تكيفت في الحياة في المناطق البحرية المفتوحة وجدت في مساحات أكثر إتساعاً بكثير من الحيتان المتقدمة عنها. حفرياتهم وجدت بعيدة جداً عن جنوب آسيا، فوجدت فيما يعرف حالياً بجورجيا، لويزيانا وكولومبيا البريطانية.
خلال عصر الأيوسين، كانت معظم الأماكن التي وجدت بيها الحيتان المتأخرة متقاربة إلي حد كبير من بعضها. في الحقيقة، معظمهم كان علي حافة بحر قديم في هذا العصر يسمي Tethys، ويعد البحر المتوسط، بحر قزوين، البحر الأسود وبحر آرال من بقاياه.
التوزيع الجغرافي-الحيوي (biogeographic) للحفريات يتوافق مع النمط التي تفرضه نظرية التطور للحيتان؛ ففي البداية الحيتان تواجدوا في مساحات جغرافية صغيرة ومحدودة، ولم تصبح منتشرة حول العالم حتي تطوروا للحياة في الماء كحيوانات مائية ولم يعد لهم أي إرتباط بالأرض.
10- دليل الترتيب الزمني (Chronological evidence)
الحلقة الأخيرة من شروحنا للأدلة التي نستند عليها في تحقيق تطور الحوت من الثدييات الأرضية تأتي من طرح سؤال أساسي (لماذا تطورت الحيتان من الأصل في الوقت التي تفترضه نظرية التطور تحديداً ؟). التطور بشكل عام هو نتيجة للتغيرات البيئية والفرص المتاحة للأنواع للتكيف معها. في بداية عصر السينوزويك (من 65 مليون سنة تقريباً) أتيحت للثدييات علي الأرض فرص للإنتشار والتنوع، ويرجع هذا بشكل جزئي إلي الفراغ علي الكوكب بسبب عمليات الإنقراضالتي حدثت في نهاية فترة فترة ال Cretaceous وقبل عصر ال Cenozoic. وبسبب أن الزواحف لم تعد مسيطرة وحاكمة للأرض، نتج عن ذلك وللمرة الأولي فرص أسهل للثدييات في الحياة.
قبل فترة الإنقراض الهائلة للأنواع التي حدثت في نهاية فترة الCretaceous، كانت الزواحف المائية في عصر الMesozoic (أكثر من 65 مليون سنة) كالplesiosaurs, ichthyosaurs, mosasaursوالتماسيح المائية في الأغلب تتغذي علي أي كائن من الثدييات يقترب من الشواطئ بحثاً عن غذاء.
وبإختفاء هذه الضواري، التطور وبسرعة أنتج ثدييات جديدة ومنها الحيتان في طريقها لنمط حياة مائية بشكل كامل. التحول إستغرق تقريباً 10-15 مليون سنة لينتج حيتان مائية تعيش في الأعماق ولها القدرة علي السماع الموجه تحت الماء. عملية التطور تفترض أن الحيتان لم تكن ببساطة أن تظهر أو تنتشر قبل الأيوسين، وأن الثدييات يفترض بها أن تنتشر إلي بيئات مائية كما حدث ذلك أيضاً إلي بيئات أخري تفرغت من الحياة التي سيطرت علي الزواحف بنهاية الCretaceousوهو ما نراه بوضوح في السجل الحفري لهذه الفترات الزمنية.
إستنتاج وحيد نخرج به بعد شرح كل الأدلة السابقة (الحيتان تطورت من ثدييات أرضية). رأينا أنه هناك عشر مجالات مستقلة من الدراسة تطرح الدليل علي أن الحيتان لها سلف مشترك مع ذوات الحوافر. لا يوجد أي طرح آخر له من القاعدة العلمية الصلبة يشرح "كيف" ظهرت الحيتان للحياة سوي ما تطرحه نظرية التطور من أن الحيتان تطورت من ثدييات أرضية.
وبعد هذا كله لا يكون لطرح معارضي التطور عن إستحالة كون ثدييات أرضية مشعرة تطورت إلي حيتان تعيش تحت الماء أي أرضية صلبة أمام الدليل المستدل عليه من عدة مسارات مختلفة ومتوازية من الدراسات العلمية.
هكذا يعمل العلم ولهذا أنا أصدق العلم ..
ملحق للمقالة ..
ذكرنا ونحن نشرح الدليل الحيوي، أن أحدث الدراسات الجزيئية وضعت كائن الHippo كأقرب الأنواع للحيتان الحديثة بل وأقرب من الثدييات ذوات الحوافر الartiodactyls(الحالية). هذه النتائج لم تغير من الحقيقة التي تطرحها نظرية التطور عن تطور الحيتان الحديثة من ثدييات أرضية وتحديداً الartiodactyls وشرحنا الأدلة عليها في هذه المقالة.
التصور الجديد أن الartiodactylsتفرعت منها مسارين تطورين أحدهما يصل بنا إلي الحيتان الحديثة –وهي الحالة التي شرحناها في هذا المقال- والمسار الثاني يصل بنا إلي الHippo مروراً بالanthracotheres وهو سلف الHippoالأرضي وعاش من 15 مليون سنة تقريباً؛ ويعود السلف المشترك للحيتان الحديثة والHippo إلي 55 مليون سنة تقريباً–كائن أرضي أقرب منه مائي-ثم السلف من الartiodactylsالذي تفرع منه كل الحيوانات ذوات الحوافرartiodactyls الحديثة والحيتان والHippos من65 مليون سنة تقريباً.
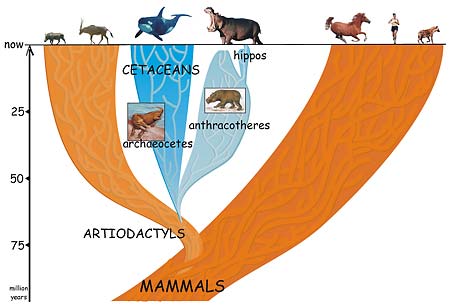
(Credit: Jean-Renaud Boisserie/UC Berkeley)
المصادر
(1) المقالة التي شرحناها هنا هي في الأصل مقالة بالإنجليزية منشورة علي موقع talkorigins بعنوان (The Origin of Whales and the Power of Independent Evidence) وهي تكون أغلب أجزاء المقالة مع إضافات أخري.
http://www.talkorigins.org/features/whales/
(4) http://www.talkorigins.org/faqs/comdesc/section2.html#fig2.2.2